- 品牌
- 有梦生物
- 服务项目
- 扫描
1. 生物学中的全景扫描是整合显微成像、光谱分析与计算机算法的前沿技术,能对生物样本进行全域高精度观测,其分辨率可达纳米级,从单细胞的细胞器结构到完整组织切片的细胞排列,都能清晰捕捉细微结构与动态变化。例如在追踪胚胎发育中细胞迁移轨迹时,可连续数小时实时记录,结合荧光标记精细定位蛋白质在细胞内的分布与转运过程,为细胞生物学中细胞分化、信号传导等研究提供三维全景数据,极大推动了对生命活动微观机制的深入理解,帮助科研人员发现了多种此前未被观测到的细胞间相互作用模式。全景扫描观察染色体联会,分析减数分裂中同源染色体的配对过程。福建芯片全景扫描电话多少
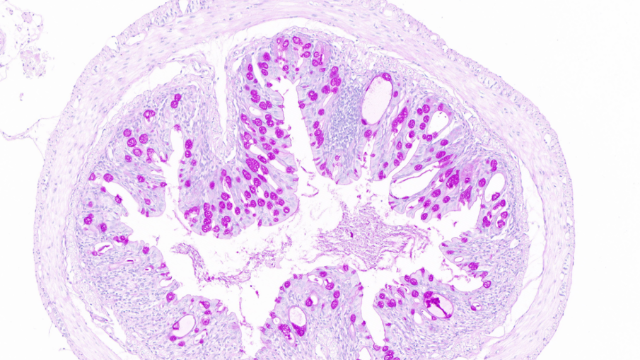
在血管生物学研究中,全景扫描技术 通过多模态动态成像系统,实现了对血管网络 发生-重塑-病理演变 全过程的 四维可视化解析(三维空间+时间维度)。该技术整合 双光子***显微术(2P-LSM)、光片荧光显微镜(LSFM)和 超声微血流成像,可在单细胞精度追踪:血管新生机制转基因斑马鱼模型 的全景扫描显示,VEGF-A165 诱导的 内皮前列细胞 以 "丝状伪足探路" 方式(延伸速度3μm/min)引导血管定向生长超分辨显微镜(dSTORM)发现 Notch1-Dll4信号轴 通过调控内皮细胞 核内Hes1蛋白振荡频率(每90分钟1次)决定血管分支间距**血管异常性全***透明化扫描 揭示**血管存在 "盲端-环状-螺旋" 三种畸形构型,其 壁细胞覆盖率 不足30%(正常血管>70%)量子点标记血流成像 显示**血管通透性增加100倍,导致 "血浆渗漏-间质高压" 恶性循环***靶点发现药物响应全景扫描平台 证实,抗VEGFR2纳米颗粒能选择性阻断 直径<15μm 的新生血管,使**灌注量下降80%单细胞转录组耦合成像 发现 SEMA3E-PlexinD1 通路是***中 血管钙化 的关键开关河南荧光单标全景扫描售价用全景扫描研究蚂蚁导航,观察其利用视觉标记识别路径的行为。
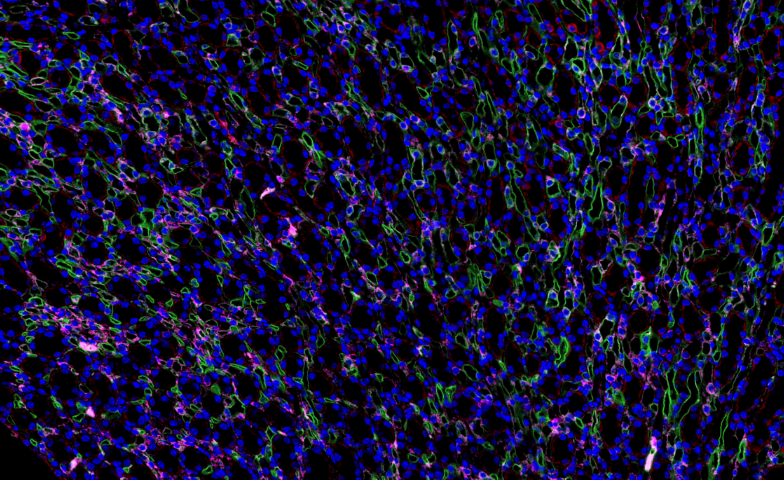
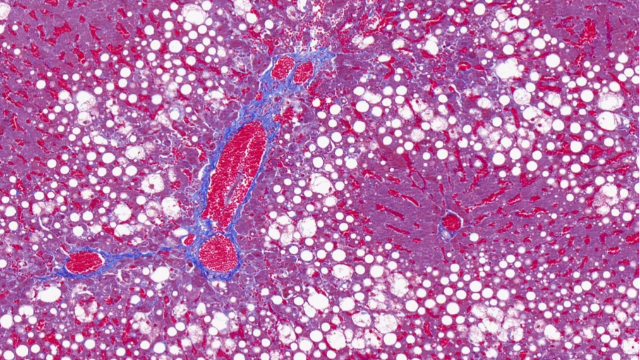
在植物光合作用研究中,全景扫描技术 通过多尺度成像与功能分析联用,系统揭示了 光合结构-功能耦合机制。该技术整合 冷冻电镜断层扫描(Cryo-ET)、荧光寿命成像(FLIM)和 原子力显微镜(AFM),实现了从 类囊体基粒堆叠(单层厚度10-12nm)到 全叶光合活性 的跨维度解析。以高光胁迫(1500μmol·m⁻²·s⁻¹)研究为例:超微结构层面:冷冻电镜全景扫描 显示PSII超复合体在强光下2小时内发生 二聚体解离(从80%降至35%)类囊体膜出现穿孔(直径50-100nm),伴随 Cyt b6f复合体空间重排生理动态层面:多光谱荧光扫描 捕获到叶黄素循环(VDE酶***)在5分钟内启动,非光化学淬灭(NPQ)效率提升3倍拉曼成像 发现β-胡萝卜素在强光区优先降解(1530cm⁻¹特征峰减弱60%)分子调控层面:原位杂交全景扫描 显示 PsbS基因 在束鞘细胞中表达量激增8倍,与抗光氧化关键蛋白(如PTOX)共定位
0. ***。,学研究中,全景扫描技术用于观察***的菌丝网络结构、孢子形成及与其他生物的共生关系,通过成像系统扫描***在培养基或自然环境中的生长状态,分析菌丝的分支模式、长度及分布特征。结合代谢产物分析,揭示***的代谢功能及与植物、微生物的相互作用,例如在菌根***研究中,发现了***菌丝与植物根系的紧密结合及养分交换的路径,为提高植物的养分吸收能力和抗逆性提供了依据,同时也有助于开发***来源的生物农药和生物肥料。全景扫描分析肌肉干细胞,呈现其在肌肉损伤后的**与分化。
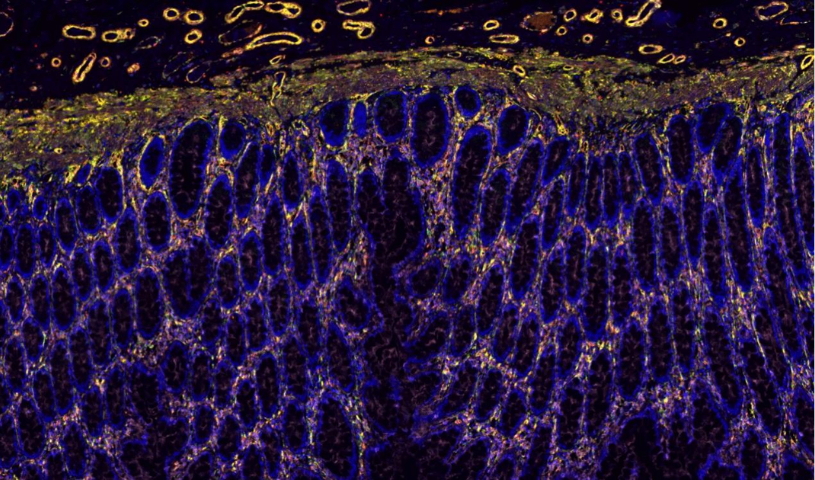
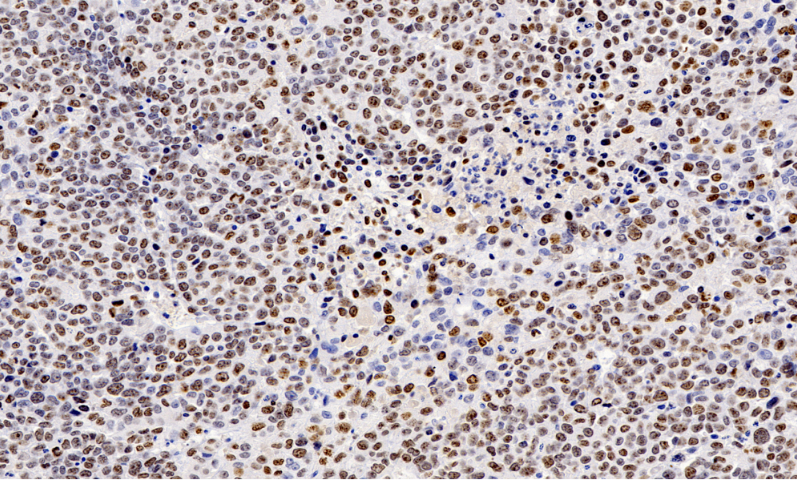
0. 免疫学研究中,全景扫描技术可对免疫***如淋巴结、脾脏进行全域成像,清晰呈现 T 细胞、B 细胞、巨噬细胞等免疫细胞的空间分布及相互作用。通过标记不同免疫细胞表面的特异性分子,结合实时成像,能追踪免疫细胞在抗原刺激后的活化、增殖及迁移轨迹,揭示免疫应答的启动与调控机制。例如在研究自身免疫性疾病时,全景扫描发现了免疫细胞异常聚集与组织损伤的关联模式,为疾病的免疫调节***提供了新的靶点和策略,同时也助力疫苗免疫效果的评估,通过观察免疫细胞的活化程度判断疫苗的有效性。全景扫描观察鞭毛运动,揭示细菌借助鞭毛实现定向移动的机制。黑龙江荧光双标全景扫描性价比
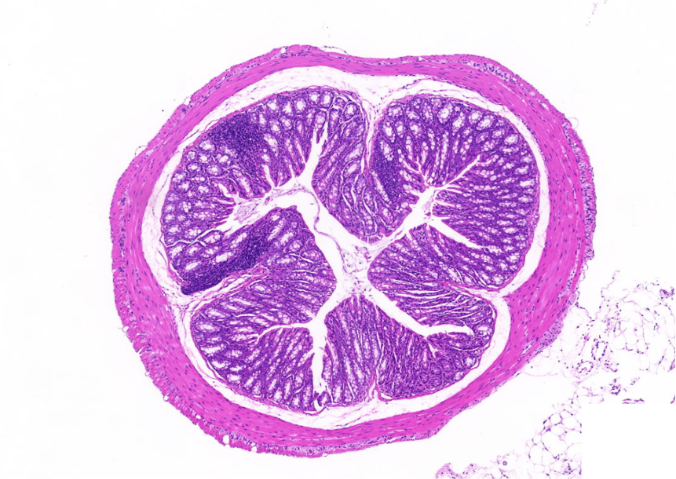
用全景扫描研究蛙类**,呈现蝌蚪尾部消失与四肢形成的过程。福建芯片全景扫描电话多少
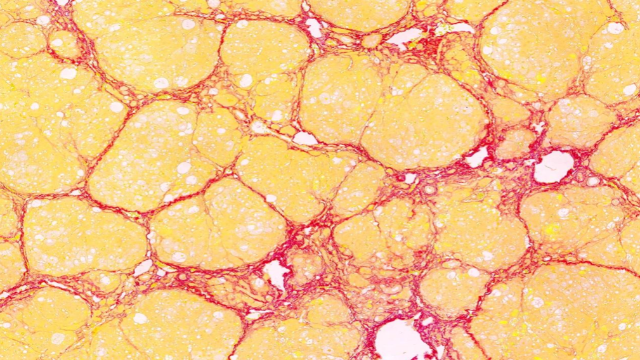
0. 全景扫描技术在生物力学研究中用于分析生物材料的力学性能与结构的关系,通过力学测试与成像技术结合,扫描骨骼、肌腱、软骨等生物组织的微观结构,测量其在受力情况下的变形、应力分布等力学参数。结合计算机模拟,揭示生物材料的力学适应机制,例如在研究骨骼的结构与强度关系时,全景扫描发现了骨骼内部的孔隙结构、纤维排列与骨骼承重能力的关联,为开发仿生材料和骨科植入物提供了设计依据,同时也有助于理解运动损伤的发生机制和康复***的原理。福建芯片全景扫描电话多少
- 四川PAS染色全景扫描 2026-01-01
- 上海荧光双标全景扫描销售电话 2026-01-01
- 上海荧光全景扫描性价比 2026-01-01
- 江西髓鞘全景扫描大概价格 2025-12-31
- 山东荧光三标全景扫描大概价格 2025-12-30
- 吉林刚果红染色全景扫描咨询报价 2025-12-30
- 黑龙江免疫组化全景扫描市场价格 2025-12-30
- 北京油红O全景扫描大概多少钱 2025-12-29
- 山东TRAP染色全景扫描咨询报价 2025-12-29
- 广西芯片全景扫描性价比 2025-12-29
- 上海全景扫描市场价格 2025-12-29
- 广西荧光单标全景扫描欢迎选购 2025-12-26