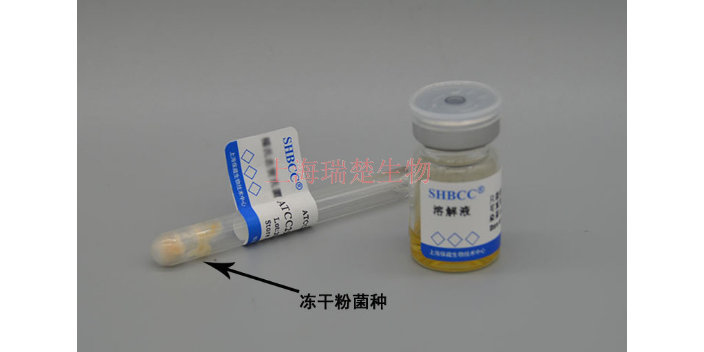
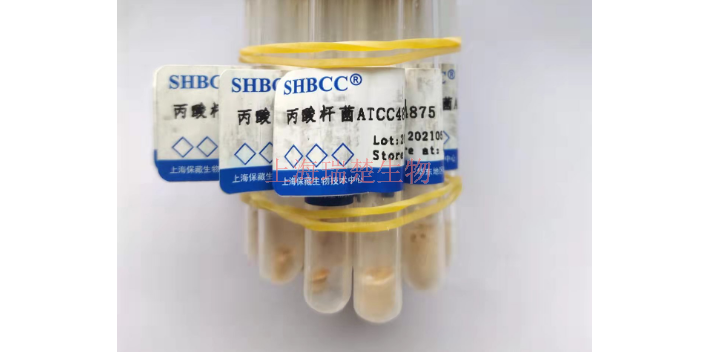
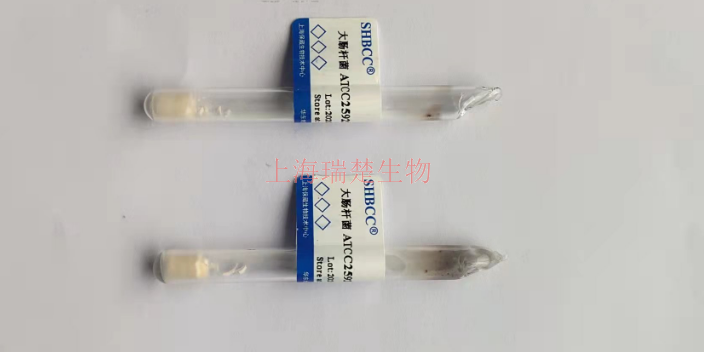
- 品牌
- 上海保藏生物技术中心 SHBCC
- 贮藏
- 4-10度冷藏
- 生产企业
- 瑞楚生物科技(江苏)有限公司
- 规格
- 冻干管
- 产地
- 江苏盐城
- 厂家
- 上海保藏生物技术中心
- 有效期
- 36个月
- 运输条件
- 冰袋加顺丰快递
- 长期储存温度
- 4-10度
- 活化之后储存条件
- 4-10度
- 转接时间
- 3个月转接一次
- 长期储存方法
- 甘油菌-80度可以保存2年以上
- 活化方法
- 好氧菌斜面活化,厌氧菌培养皿厌氧培养,兼性厌氧液体培养基培养
- 打管方法
- 直接敲开
- 接种量
- 活化时取0.2-0.3ml溶解液后全部溶解接种到培养基上
- 活化代数
- 活化3代活力比较好
- 操作硬件设施要求
- 二级生物安全柜内操作
除了上述提到的因素,还有一些其他因素可能影响枯草芽孢杆菌芽孢的存活时间:1.**氧气浓度**:芽孢在缺氧条件下通常更容易存活。高氧环境可能会加速芽孢的老化和失活。2.**湿度**:湿度对芽孢的存活和萌发有影响。高湿度环境可能促进芽孢的萌发,而低湿度环境则有助于芽孢的长期保存。3.**光照**:紫外线辐射对芽孢有破坏作用,尤其是在波长较短的UV-C范围内。光照强度和暴露时间都会影响芽孢的存活。4.**营养基质**:芽孢在不同的营养基质中形成的芽孢可能具有不同的热抗性和化学抗性。例如,某些培养基中形成的芽孢可能对热处理更敏感。5.**微生物代谢产物**:芽孢在形成过程中产生的代谢产物,如吡啶二羧酸(DPA)和钙离子(Ca2+),在芽孢的热抗性中起关键作用。这些代谢产物的含量和比例可能影响芽孢的存活时间。6.**物理损伤**:机械损伤如振动、冲击等可能会破坏芽孢的结构,降低其存活率。7.**化学污染**:某些化学物质如消毒剂、清洁剂等可能会对芽孢产生毒性作用,影响其存活。8.**基因因素**:不同菌株的芽孢可能具有不同的基因型,这会影响其对环境压力的响应和存活能力。基因突变或基因表达的差异可能导致芽孢的热抗性和其他抗性特性的变化。居海绵华美菌,这种细菌是从大韩民国济州沿海地区的海洋海绵中分离出来的。乳肠球菌菌种

热生泛菌(Pantoea)是一种具有独特特性的微生物,以下是其主要特点和介绍:1.**形态特征**:-热生泛菌的菌体呈杆状,革兰氏阴性。菌落2-3mm,不规则圆形,微黄,有光泽,表面隆起,雪花状,有褶皱,不透明,边缘不整齐。2.**生长条件**:-热生泛菌属于好氧或兼性厌氧发酵型革兰氏阴性杆菌,合适温度为30℃。D-葡萄糖和其他糖类可产酸,但不产气。氧化酶阴性,接触酶阳性,吲哚阴性,M-R可变。不产生H2S,不水解脲素。大多数菌株可生长于KCN,还原硝酸盐。3.**代谢特性**:-热生泛菌具有代谢和发酵类型的化能异养菌。赖氨酸和鸟氨酸脱羧酶以及精氨酸双水解酶皆阴性(Gavini等发现30%的成团泛菌鸟氨酸脱羧酶是阳性)。丙二酸盐利用在菌株间可变。4.**应用价值**:-热生泛菌的主要用途为研究,具体用途包括酿造白酒和产糖化酶。5.**环境分布**:-热生泛菌分离自植物表面、种子、土壤和水,也可从动物和人的伤口、血和尿中分离到。6.**生物危害程度**:-热生泛菌是人的条件致病菌。这些特点使得热生泛菌在微生物学研究和应用领域中具有重要的价值。结节青霉菌种蓝色小单孢菌在甘油天冬素琼脂上不生长,而在无机盐淀粉琼脂上无孢子层形成 。但在纤维素上生长良好 。
拉氏根瘤菌(Rhizobiumleguminosarum)主要与豆科植物形成共生关系,进行固氮作用。其与非豆科植物共生的可能性相对较低,主要原因如下:1.**宿主专一性**:拉氏根瘤菌对豆科植物具有较高的宿主专一性,它们通过识别豆科植物释放的特定信号分子(如黄酮类化合物)来触发共生信号的交流。这种专一性使得它们很难与非豆科植物建立有效的共生关系。2.**信号交流障碍**:非豆科植物可能不产生或产生较少的根瘤菌所需的信号分子,导致根瘤菌无法识别并响应这些植物的信号,从而阻碍了共生关系的形成。3.**根瘤形成机制**:拉氏根瘤菌与豆科植物形成根瘤的机制是高度特化的。这种机制涉及到复杂的分子和细胞水平的相互作用,包括植物根部的细胞分化和根瘤菌的入侵。非豆科植物可能缺乏这种特化的机制,使得根瘤的形成变得困难。4.**固氮酶活性**:拉氏根瘤菌的固氮酶系统是针对豆科植物的共生固氮需求而优化的。这种系统在非豆科植物中可能无法有效发挥作用,因为不同植物对氮素的需求和利用方式可能存在差异。5.**生态位竞争**:在自然环境中,拉氏根瘤菌可能面临与其他微生物(如其他根瘤菌或非根瘤固氮菌)的竞争。
伊氏副球菌(Paracoccusisoporae)是一种属于副球菌属(Paracoccus)的微生物,具有以下特点:1.**形态特征**:-伊氏副球菌的菌体呈球形或近球形,单个、成对或成簇排列,革兰氏阴性。2.**生长条件**:-伊氏副球菌是好氧或兼性厌氧非发酵革兰氏阴性杆菌。在好氧条件下进行呼吸代谢,当硝酸盐、亚硝酸盐或氧化氮存在时,能以它们为电子受体营厌氧生长。3.**代谢特性**:-在厌氧条件下,伊氏副球菌能够还原硝酸盐到亚硝酸盐、氧化氮和氮气(N2)。有些种类在H2和CO2存在时可以进行自养生长,或用各种有机化合物作为碳源进行异养生长。4.**主要价值**:-伊氏副球菌的主要用途为研究,具体用途包括发酵普洱茶。5.**环境分布**:-伊氏副球菌出现在土壤、天然和人工的盐水中。6.**生理特性**:-伊氏副球菌的合适生长温度为25~30℃,氧化酶和接触酶皆阳性。这些特点使得伊氏副球菌在微生物学研究和应用领域具有重要的价值。居海绵华美菌的培养条件为DSMMedium 514,在28°C的条件下培养,可以使用BACTO MARINE BROTH 作为培养基 。
千叶类芽胞杆菌在土壤修复过程中可能会遇到的挑战以及克服方法主要包括:1.**重金属有效态含量的提高**:千叶类芽胞杆菌能够通过自身的代谢活动降低土壤pH值,从而增加土壤中重金属的有效态含量。这可能会提高植物对重金属的吸收,但也可能导致重金属毒性增加。2.**土壤酶活性的影响**:千叶类芽胞杆菌的加入可能会影响土壤中酶的活性,这对于土壤生态系统的健康和功能至关重要。研究显示,芽孢杆菌能够提高土壤磷酸酶、脲酶和蔗糖酶的活性。3.**植物抗逆性的提高**:在重金属胁迫下,千叶类芽胞杆菌可以通过提高植物的抗氧化酶活性,如过氧化物酶(POD)、超氧化物歧化酶(SOD)和过氧化氢酶(CAT),来增强植物的抗逆性。4.**植物生长促进**:千叶类芽胞杆菌可以促进植物生长,提高其生物量,这对于植物在修复过程中吸收更多重金属至关重要。5.**微生物与植物的协同作用**:构建微生物与植物的联合修复系统可以提高土壤修复效率。千叶类芽胞杆菌与植物的联合修复体系,可以更有效地活化土壤中的重金属,并促进植物对其的吸收。在MRS培养基上菌落呈圆形、白色,凸起,表面光滑、湿润,边缘整齐。 兼性厌氧细菌,生长温度范围2~53℃。食苯芽孢杆菌菌种
长野解支链淀粉芽孢杆菌能够产生一种耐热的酸性普鲁兰酶(E.C.3.2.1.41),这种酶能够分解支链淀粉 。乳肠球菌菌种
巴塞尔贪铜菌(Cupriavidusbasilensis)在生物修复重金属污染方面具有潜在的应用,尽管搜索结果中没有直接提及该菌种具体的生物修复机制。然而,基于其特性和β-变形菌纲的一些共性,我们可以推断其可能的生物修复机制:1.**重金属耐受性**:巴塞尔贪铜菌可能具有耐受多种重金属的能力,这使得它能够在重金属污染的环境中生存并发挥作用。2.**生物吸附**:该菌可能通过细胞表面的官能团与重金属离子形成稳定的复合物,从而吸收和固定重金属离子,减少其在环境中的迁移性和生物有效性。3.**生物转化**:巴塞尔贪铜菌可能具有将重金属转化为较低毒性形态的能力,例如将六价铬还原为三价铬,或将有机砷氧化为无机砷等。4.**植物-微生物联合修复**:巴塞尔贪铜菌可能与超积累植物形成共生关系,通过分泌植物生长调节物质和有机配位体,促进植物对重金属的吸收和转运,提高植物修复的效率。5.**分泌有机酸**:该菌可能通过分泌有机酸(如柠檬酸、琥珀酸等)与重金属离子络合,改变土壤中重金属的存在形态,降低其毒性并促进植物吸收。6.**产生铁载体**:巴塞尔贪铜菌可能通过产生铁载体与重金属离子络合,减少宿主植物对重金属的吸收,从而减轻重金属对植物的危害作用。乳肠球菌菌种


- 球孢链霉菌七叶树变种菌种 2024-12-23
- 五原链霉菌菌株 2024-12-23
- 蠕虫生节丛孢菌种 2024-12-23
- 噬糖盐红菌 2024-12-23
- 福氏志贺氏菌 2024-12-23
- 巴伐利亚鸟氨酸芽孢杆菌菌株 2024-12-23
- 肠沙门氏菌肠亚种赛罗血清型 2024-12-23
- 裸胞壳属 2024-12-23